Respiratory structure and gas exchange
The cardiac and respiratory structures are contained within the Thoracic cavity, figure1. The respiratory system's primary function is to supply the blood with oxygen (O2) so that it can deliver O2 to the body. The respiratory system does this through breathing. When we breathe, we inhale air to obtain oxygen and exhale to excrete carbon dioxide (CO2).
As each lung grows it develops an outer layer; the visceral pleura. Outside this the parietal pleura develops, which attaches to the thoracic wall. Between them is a tiny, almost non-existent space, called the pleural cavity, filled with serous fluid, creating enough surface tension to act as glue, Figure 2 shows the arrangement.
Thus as the parietal pleura is drawn out by the thoracic chestwall, the visceral pleura moves with it, forcing the lung tissue to expand..
Figure 3, shows the air entry route through the mouth and the nose, larynx and trachea - the tube into the thoracic cavity, where it splits into the two bronchi.
 Reference ranges for blood tests, showing adult ma...
Reference ranges for blood tests, showing adult ma... blood
blood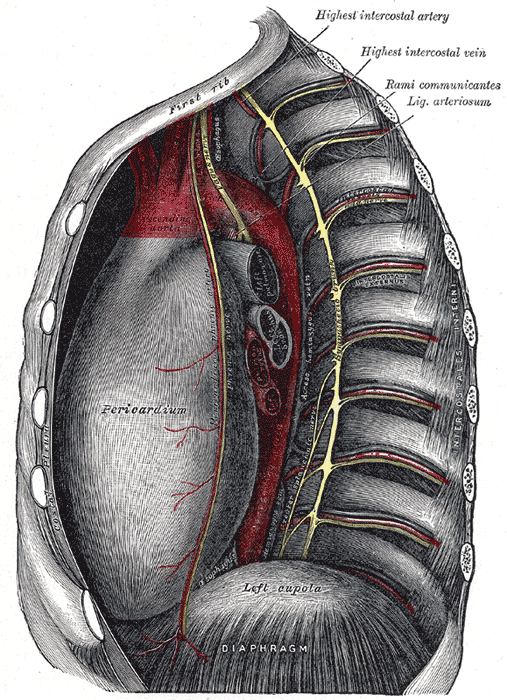 Thoracic cavity seen from left. Lungs, among other...
Thoracic cavity seen from left. Lungs, among other...Each bronchus then divides again forming the bronchial tubes, which lead directly into the lungs dividing into many smaller tubes, connecting to tiny sacs called alveoli. The average adult's lungs contain about 600 million of these spongy, air-filled sacs that are surrounded by capillaries (figure 4), this arrangement produces an enormous surface are for gas exchange; oxygen diffuses through the alveoli and capillary walls and then into the arterial blood, figure 5.
CO2 leaves and O2 enters the blood due to the partial pressure gradient between the alveolar space and the alveolar capillary, figure 5, moving by diffusion. With the enormous surface area; two-way gaseous transfer takes about one second.
When blood first arrives at the pulmonary capillary at its arteriole end, the partial pressures of CO2 and oxygen are:
PCO2 = 45 mmHg
PO2 = 40 mmHg
There's a lot of CO2 in blood since all the waste CO2 from the tissues of the body is in it. The partial pressure of CO2 in the atmosphere is much tinier than this, <0.3 mmHg.
The partial pressure of O2 is only 40 mmHg in this blood; because of the O2 given-up to bodily tissues during circulation. The Fick Equation;
VO2 = Q x aO2 - vO2, aO2 = arterial circulation O2, vO2 = venous circulation O2
Is an expression of the O2 lost due to cellular respiration needs through increased exercise. The partial pressure of oxygen in the atmosphere was 160 mmHg, which is much higher than 40 mmHg!
In this time the blood becomes nearly 100% saturated with O2 and loses its excess CO2. Meanwhile, the waste-rich blood from the veins releases its CO2 into the alveoli. The CO2 follows the same path out of the lungs when you exhale.
Table 1; Factors affecting alveolar to blood gas transfer
This transfer in both directions can be affected as follows;
-Surface area
çDecreased in disease, e.g. emphysema
çMore capillaries open during exercise, increasing area available
-Barrier thickness
çSome disease thickens the one cell thickness of the capillaries reducing exchange speed
çDiffusion co-efficient can be changed biochemically to reduce exchange speed
-CO2 is 20 times as soluble as O2; means alveolus can be smaller using O2
çFacilitates gas exchange in the correct direction!
These gaseous movements are driven through the change in interalveolar pressure resulting from the change in lung volume (Boyle's law) accompanying diaphragm contraction and relaxation, figure 6.
The intrapleural pressure isn't the same throughout the lungs; more negative at the top. The alveoli at the top of the lungs inflate first. In exercise the lower alveoli also inflate. At this stage another control mechanism occurs, the Hering-Breuer response. In this the medulla's rhythm is supplemented by stretch receptors in alveoli, bronchioles and bronchi. They sense the alveoli reaching maximum inflation and send a message to the medulla to exhale to prevent over inflation.
Additionally gas exchange at the lungs is matched to respiratory needs partly by dilating or contracting smooth muscle in the openings to the alveoli under control of the centres in the medulla.
This dead space fresh air is pushed out during the following expiration and is replaced by breath containing a high amount of CO2. On the next breath this alveolar gas from the dead space is drawn back into the alveoli before the incoming fresh air.
During inhalation - always an active process, the intercostal muscles and the diaphragm contract. The volume of the lungs increases as the diaphragm moves down and the rib cage expands, up and out; the pressure in the lungs falls, air rushes in, figure 7.
Resting inhalation relies mainly on the diaphragm, but during exercise the action of the external intercostal muscles becomes the most important factor. In forced inhalation during maximal exercise the Sternocleidomastoid, Platysma, and the three-scalenus muscles in the neck also help raise the ribs.
At rest expiration is passive; lungs contract due to elasticity, and the surface tension of the alveoli (from surfactant); the intercostals and diaphragm relax, and the forces of recoil restore the thoracic cavity's smaller volume (Recoil due to LaPlace's law; P = 2T / r). Thus pressure becomes greater in lungs than in the atmosphere and air rushes out.
During forced exhalation, as in maximal exercise, the internal intercostals pull the ribs downward and inward; the latissimus dorsi and quadratus lumborum assist, and the abdominal muscles contract forcing the diaphragm up, reducing the thoracic cavity.
The changes in intra-abdominal and intrathoracic pressure that accompany forced breathing also assist in returning venous blood back to the heart, essential in maximal exercise as it reduces the blood pressure increase required.
The respiratory centre is grey-matter in the pons and the upper Medulla; responsible for rhythmic respiration, this centre can be divided into an inspiratory and an expiratory centre in the Medulla, an apneustic centre in the lower and midpons and a pneumotaxic centre in the rostral-most part of the pons. This respiratory centre is very sensitive to the pCO2 in the arteries and to the pH level of the blood, triggering the breathing process so that more O2 can enter the body to replace the O2 that has been utilised. The Medulla Oblongata sends neural impulses down through the spinal chord and into the diaphragm.
Blood is a sophisticated system for delivery of O2 sufficient for the body's needs. A sedentary person consumes 600L O2 and produces 480L of CO2 daily; this increases substantially with activity, as shown in the experimental results given in appendix 1. Dissolved O2 in blood plasma would supply only 10L daily. The solubility of O2 is low, at human body temperature (37oC) only 0.46cm3 dissolving in 100cm3 of water/plasma. An important oxygen binding protein, haemoglobin, evolved so that aerobic metabolic processes were no longer limited by the solubility of O2 by directly transporting oxygen in the red blood cells (erythrocytes).
Haemoglobin increases the capacity of blood to transport O2 by about 60-fold and has the ability to combine readily with O2 where its concentration is high, i.e. at the respiratory surface, and to release it readily where its concentration is low i.e. in the tissues.
Haemoglobin is a tetramer composed of 4 polypeptide chains, figure 13, 2 identical a chains and 2 b chains. Each chain contains haem, a porphyrin ring system with an iron ion (Fe2+) to which oxygen binds; four O2 in total.
Its ability to bind O2 lies in 3-D conformational changes resulting from one subunit binding oxygen, which brings about a change in properties of the three remaining subunits so that they are able to bind oxygen more strongly. This influence of the binding of O2 by one subunit on the O2 affinity of the other subunits is called cooperativity. This change is from the taut (T) state, to the relaxed (R) state, figure 14.
The affinity for the fourth oxygen to bind is approximately 300 times that for the first. This behaviour results in an oxygen dissociation curve (a plot of saturation of oxygen-binding sites vs. partial pressure of oxygen) that is sigmoidal, rather than hyperbolic. Hyperbolic curves are exhibited by monomeric molecules such as myoglobin, figure 15, which transports oxygen within cells to the mitochondria once it's diffused into them from the blood.
The significance of the sigmoidal curve is haemoglobin becomes highly saturated at high PO2 levels and releases a significant amount of O2 at low PO2 pressures. Thus haemoglobin becomes more efficient at O2 delivery under maximal exercise conditions and is helped in this by the Bohr shift. The Bohr shift helps the body release more O2 to respiring tissues when the pH is more acidic. Thus, the oxygen debt that builds up through the production of lactate both stimulates the medulla to increase breathing rate and instigates the Bohr shift, inducing haemoglobin to release more oxygen to working tissues.
CO2 produced by body tissues diffuses into the interstitial fluid, then blood plasma. <10% remains in the plasma as dissolved CO2. ~70% diffuses into erythrocytes, where ~20% binds with haemoglobin to form carboxyhaemoglobin. ~50% reacts with H20 in the erythrocytes to form carbonic acid, which is immediately broken-down by carbonic hydrase into bicarbonate and H+, as a result the change in blood pH is only a few hundredths of a pH unit, figure 16. This is reversed in the lungs so that H2O and CO2 are exhaled. The reversibility of the carbonic acid-bicarbonate conversion also helps buffer the blood depending on the H+ or pH level triggering chemoreceptors in the aorta, carotid bodies or central receptors in the medulla. Chlorine goes into the red blood cells when bicarbonate comes out. This is referred to as the chloride shift.
The inside layer of arteries and veins, figure 17, is very smooth, allowing the blood to flow quickly down the tube space; lumen, reducing peripheral resistance to blood flow. The outside layer of an artery is muscular and very strong, allowing the blood to flow forcefully. Both arterial and venous smooth muscle is under sympathetic nervous control form the medulla.
Table 2; Cardiac Terminology relating to sports and exercise
Cardiac cycle.The cycle has two phases:Diastolic - the relaxed phase when blood flows into the heart's atria form the vena cavae and the pulmonary vein.Systolic - the contract phase when blood is pumped in the aorta and pulmonary artery.Blood pressure measurements are an expression of the Systolic over the diastolic pressures, normal 120/80 mmHg. One cycle is roughly 62% diastolic and 38% systolic.
Stroke Volumeis a measure of the amount of blood pumped per beat (contraction), i.e. out of the left ventricle into the aorta. It's measured by deducting the End stroke Volume (ESV) from the End diastolic volume (EDV).SV = EDV - ESVEDV is the volume of blood left in the left ventricle at the end of the diastolic phase. ESV is the volume of blood left in left ventricle at the end of the systolic phase. Normal SV ~ 60-80ml
Ejection Fraction (EF%)is a measure of the percentage of blood ejected by the left ventricle compared to the left ventricles original volume. It's expressed as;(SV/EDV) x 100 = EF%
Cardiac Output (Q)is the amount of blood pumped by the heart in one (1) minute. It's calculated by multiplying the stroke volume by the heart rate in beats per minute (bpm);HR (bpm) x SV = Q
Q increases with exercise as the heart rate increases proportionate to the intensity of the exercise until maximal heart rate is reached. Stroke volume also increases proportionately up to a point of 60% os maximal heart rate, table three summarises the long and short-term effects of maximal exercise on these factors.
Table 3; Short and long-term effects of exercise on cardio-circulatory system
Short-term EffectsLong-term effects
Heart rate goes up; HR ÃÂHeart rate goes down; HR ï
Blood pressure goes up; BP ÃÂBlood pressure goes down; BP ï
Stroke Volume goes up; SV ÃÂStroke Volume goes up; SV ÃÂ because the heart is able to pump more with each individual stroke
Cardiac Output (HR x SV) goes up; CO ÃÂCardiac Output - not measured as one goes up the other down so not a meaningful long-term measure
Body Temperature increase; BT ÃÂ
Blood flow is re-directed to the areas working, e.g. muscles
The delivery of O2 and removal of CO2 is more efficientThe delivery of O2 and removal of CO2 is more efficient
The rate of breathing increases
The heart gets bigger and stronger
The Cardiac System Structure and Function
Table 4: Typical heart rates
ActivityBeats per Minute (bpm)
Sleeping60 - 70
Walking80 - 100
Riding a bike110 - 130
After a Meal80 - 90
Watching an exciting sports event80 - 90
Moderate fever90
The Heart, figure 18, is made up of powerful cardiac muscle; Myocardium, packed with mitochondria, whose fibres contract and causing a wringing action. It really consists of two separate pumps that continuously circulate blood, roughly 5 litres in an adult human. The right atrium of the heart receives blood low in oxygen and high in CO2 from the vena cava; systemic circulation. From here blood passes into the right ventricle through a one-way valve where it's pumped into the pulmonary artery and into the lungs for alveolar gaseous exchange, figure 5. Re-oxygenated blood passes down the pulmonary vein back into the heart's left auricle, then passing into the left ventricle via another one-way valve where it's pumped into the aorta and back into the arteries of systemic circulation to begin its journey throughout the body, figure 13. In the systemic system blood flows from the heart and back to it as follows; Arteries leading to arterioles leading to capillaries leading to venules leading to veins.
Minute electrical impulses are generated by a specialised group of cells in the wall of the right atrium, called the sinoatrial (SA) node; the heart's internal, self-regulating pacemaker. It's made of specialised tissue, which spontaneously fires about once per second, ~ rest HR; 60bpm. With stimulation from certain chemicals, hormones, or exercise, the spontaneous rate can be increased to as high as 180 to 200 beats per minute. During strenuous exercise, the heart can increase stroke volume four-fold because of;
1.Frank-starling mechanism.
The ventricular walls stretch as more blood enters more blood = more stretch. Receptors in the walls detect the degree of stretch and alter ventricular contraction accordingly; the larger the stretch the more forceful the contraction.
2.Regardless of blood volume in the ventricle, the ventricle wall contracts more forcefully as you work harder.
3.Vasodilatation of the arteries occurs to reduce the peripheral resistance and thus increases the volume and velocity of blood flow.
Leaving the SA node, the impulse passes through both atria, causing them to contract and helping blood to pass into their respective ventricles. On its way to the ventricles, the impulse next encounters the AV node, another group of specialised cells.
From there, the impulse passes down an anatomic pathway called the Bundle of His, Figure 19, which divides into Left and Right Bundle Branches. The impulse moves through the Branches into the terminal Purkinje fibres, which distribute the stimulus to myocardial, cells and cause the ventricles to contract.
The electrical process that accompanies contraction is called depolarisation. The intercalated structure of the myocardium allows direct depolarisation across it, facilitating simultaneous contraction in several planes. During relaxation from the contraction, repolarisation occurs and the contracting cells return to their original state. This complex series of events occurs in less than a second, with every beat of the heart.
Extrinsic control
Two major types of nerves also travel to the heart and influence the SA node, figure 20;
qThe sympathetic nervous system from the brain and spinal cord stimulate the sinus node to fire more frequently in response to fright, flight, or fight stimulation.
qThe parasympathetic or vagal nervous system decreases heart rate or degree of contractility. The presence of hormones or chemicals such as Epinephrine and Norepinephrine (adrenalin and Noradrenalin) also can influence heart rate directly without affecting the nerves.
The (AV) node, limits the number of impulses that can pass through it in a given time period, preventing the ventricles from being driven at excessive rates. This delaying is under the control of the autonomic nervous system and hormones. At rest, under the influence of "decelerator" nerves (parasympathetic, or "vagal" nerves), inhibitory effects dominate. During exercise, "accelerator nerves" (sympathetic or adrenergic nerves) are activated, which facilitates AV node signal transmission necessary for rapid heart rates.
How would you as a sports and exercise specialist inform your public of factors to improve performance related to your knowledge of the cardio-vascular and respiratory systems?
What and how would be entirely contextual, ranging from a personal one-to-one chat about general principles involved in training, figure 21, through to detailed measures of lactate threshold, ventilation break-point and VO2max as per the results in appendix 1, in order to allow the calculation of a heart rate at 60-80% of the threshold to improve aerobic endurance by training at that level.
Very useful info
well done in explaining the basic about how the respiratory system works!
4 out of 4 people found this comment useful.