Factors of Parasitic Virulence
ABSTRACT
Why do some parasites kill the host they depend upon while
others coexist with their host? Two prime factors determine parasitic
virulence: the manner in which the parasite is transmitted, and the
evolutionary history of the parasite and its host. Parasites which
have colonized a new host species tend to be more virulent than
parasites which have coevolved with their hosts. Parasites which are
transmitted horizontally tend to be more virulent than those
transmitted vertically. It has been assumed that parasite-host
interactions inevitably evolve toward lower virulence. This is
contradicted by studies in which virulence is conserved or increases
over time. A model which encompasses the variability of parasite-host
interactions by synthesizing spatial (transmission) and temporal
(evolutionary) factors is examined. Lenski and May (1994) and Antia et
al. (1993) predict the modulation of virulence in parasite-host
systems by integrating evolutionary and transmissibility factors.
INTRODUCTION
Why do certain parasites exhibit high levels of virulence within
their host populations while others exhibit low virulence? The two
prime factors most frequently cited (Esch and Fernandez 1993, Toft et
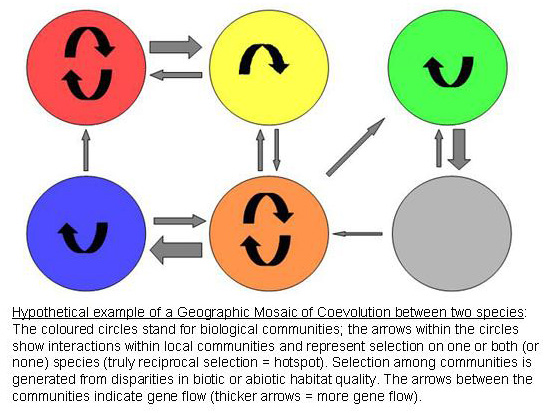 English: Graphic about the Geographic Mosaic Model...
English: Graphic about the Geographic Mosaic Model... The deer mouse (Peromyscus maniculatus) is the res...
The deer mouse (Peromyscus maniculatus) is the res... Toft Point
Toft Pointal. 1991) are evolutionary history and mode of transmission.
Incongruently evolved parasite-host associations are characterized by
high virulence, while congruent evolution may result in reduced
virulence (Toft et al. 1991). Parasites transmitted vertically (from
parent to offspring) tend to be less virulent than parasites
transmitted horizontally (between unrelated individuals of the same or
different species). Studies in which virulence is shown to increase
during parasite-host interaction, as in Ebert's (1994) experiment with
Daphnia magna, necessitate a synthesis of traditionally discrete
factors to predict a coevolutionary outcome. Authors prone to
habitually use the word decrease before the word virulence are
encouraged to replace the former with modulate, which emphasizes the
need for an inclusive, predictive paradigm for parasite-host
interaction.
Evolutionary history and mode of transmission will first be
considered separately, then integrated using an equation discussed
by Antia et al. (1993) and a model proposed by Lenski and May (1994).
Transmission is a spatial factor, defined by host density and specific
qualities of host-parasite interaction, which gives direction to the
modulation of virulence. Evolution is a temporal factor which
determines the extent of the modulation. The selective pressures of
the transmission mode act on parasite populations over evolutionary
time, favoring an equilibrium level of virulence (Lenski and May
1994).
DOES COEVOLUTION DETERMINE VIRULENCE?
Incongruent evolution is the colonization of a new host species
by a parasite. It is widely reported that such colonizations, when
successful, feature high virulence due to the lack of both evolved
host defenses and parasitic self-regulation (Esch and Fernandez 1993,
Toft et al. 1991). Unsuccessful colonizations must frequently occur
when parasites encounter hosts with adequate defenses. In Africa,
indigenous ruminants experience low virulence from Trypanosoma brucei
infection, while introduced ruminants suffer fatal infections (Esch
and Fernandez 1993). There has been no time for the new host to
develop immunity, or for the parasite to self-regulate. Virulent
colonizations may occur regularly in epizootic-enzootic cycles. Sin
Nombre virus, a hemmorhagic fever virus, was epizootic in 1993 after
the population of its primary enzootic host, Peromyscus maniculatus,
had exploded, increasing the likelihood of transmission to humans
(Childs et al. 1995). Sin Nombre exhibited unusually high mortality in
human populations (Childs et al. 1995), which were being colonized by
the parasite.
It is assumed that coevolution of parasite and host will result
in decreased virulence (Esch and Fernandez 1993, Toft et al. 1991).
Sin Nombre virus was found to infect 30.4 % of the P. maniculatus
population, exhibiting little or no virulence in the mice (Childs et
al. 1995). Similar low levels of virulence have been found in the
enzootic rodent hosts of Yersinia pestis (Gage et al. 1995). In
Australia, decreased grades of virulence of myxoma virus have been
observed in rabbit populations since the virus was introduced in 1951
(Krebs C. J. 1994). Many of the most widespread parasites exhibit low
virulence, suggesting that success in parasite suprapopulation range
and abundance may be the result of reduction in virulence over time.
Hookworms are present in the small intestines of one-fifth of the
world's human population and rarely induce mortality directly
(Hotez 1995).
Evolution toward a higher level of virulence has been regarded
as an unexplainable anomaly. Parasites which do less harm presumably
have an advantage throughout a long coevolutionary association with
their hosts. Ebert's (1994) experiment with the planktonic crustacean
Daphnia magna and its horizontally transmitted parasite Pleistophora
intestinalis suggests that coevolution does not determine the
direction of the modulation of virulence. Virulence decreased with the
geographic distance between sites of origin where the host and
parasite were collected (Ebert 1994). Thus, the parasite was
significantly more virulent in hosts it coexisted with in the wild
than it was in novel hosts. Many viruses, such as Rabies (Lyssavirus
spp.), persist in natural populations while maintaining high levels of
virulence in all potential hosts (Krebs, J. W. 1995). Extinction is
not an inevitable outcome of increased virulence (Lenski and May
1994). Increased or conserved virulence during coevolution calls
into question long held assumptions about the effect of coevolution on
parasitic virulence (Gibbons 1994). Parasitic virulence frequently
changes over coevolutionary time, but the length of parasite-host
association does not account for the virulence of the parasite.
Transmission has been identified as the factor which determines the
level of parasitic virulence (Read and Harvey 1993).
TRANSMISSION AND THE DIRECTION OF MODULATION
Herre's (1993) experiment with fig wasps (Pegoscapus spp.) and
nematodes (Parasitodiplogaster spp.) illustrates the effect of
transmission mode on parasitic virulence. When a single female wasp
inhabited a fig, all transmission of the parasite was vertical, from
the female to her offspring. The parasite's fitness was intimately
tied to the fecundity of the host upon which it had arrived. When a
fig was inhabited by several foundress wasps, horizontal transmission
between wasp families was possible. In the figs inhabited by a single
foundress wasp, Herre found that less virulent species of the nematode
were successful, while in figs containing multiple foundress wasps,
more virulent species of the nematode were successful. Greater
opportunity to find alternate hosts resulted in less penalty for
lowering host fecundity. More virulent nematodes had an adaptive
advantage when host density was high and horizontal transmission was
possible. When host density was low, nematodes which had less effect
on host fecundity ensured that offspring (i.e. future hosts) would be
available.
Low virulence is characteristic of many vertical transmission
cycles. Certain parasites avoid impairing their host's fecundity by
becoming dormant within maternal tissue. Toxocara canis larvae reside
in muscles and other somatic tissues of bitches until the 42nd to 56th
day of a 70-day gestation, when they migrate through the placenta,
entering fetal lungs where they remain until birth (Cheney and Hibler
1990). A high proportion of puppies are born with roundworm infection,
which can also be transmitted from bitch to puppy by milk (Cheney and
Hibler 1990). If host density is low, a highly evolved vertical
transmission cycle (which exhibits low virulence in the parent)
ensures the survival of the parasite population.
High virulence is characteristic of horizontal transmission
cycles. In Herre's (1993) experiment, more virulent parasites were
favored when host density was high and reduction of host fitness was
permissible. Certain parasites benefit from reduced host fitness,
particularly parasites borne by insect vectors (Esch and Fernandez
1993) and parasites whose intermediate host must be ingested by
another organism to complete the parasitic life cycle. By immobilizing
their host, heartworm (Dirofilaria immitis) and malaria (Plasmodium
spp.) increase the likelihood that mosquitoes will successfully ingest
microfilaria or gametocytes along with a blood meal. Heartworm
infestation causes pulmonary hypertension in dogs (Wise 1990),
resulting in lethargy and eventual collapse (Georgi and Georgi 1990).
Host immobility increases the opportunities for female mosquitoes to
find and feed upon hosts (Read and Harvey 1993). Infected dogs have
large numbers of D. immitis microfilaria in their circulatory systems,
again increasing the likelihood of ingestion by the insect. Many
infected dogs eventually die from heartworm, but in the process the
parasite has ensured transmission. Similar debilitating effects have
been observed in tapeworm-stickleback interaction; infected
sticklebacks must swim nearer the water's surface due to an increased
rate of oxygen consumption caused by the parasite (Keymer and Read
1991). Parasitized sticklebacks are more likely to be seen and eaten
by birds, the next host in the life cycle.
Many horizontally transmitted parasites manipulate specific
aspects of host behavior to facilitate transmission between species.
Host fitness is severely impaired in such interactions. The digenean
D. spathaceum invades the eyes of sticklebacks, increasing the
likelihood of successful predation by birds (Milinski 1990). D.
dendriticum migrate to the brains of infected ants, causing them to
uncontrollably clamp their jaws onto blades of grass, ensuring
ingestion by sheep (Esch and Fernandez 1993, Combes 1991). Infection
of a mammalian brain by rabies (Lyssavirus spp.) alters the host's
behavior, increasing the chance of conflict with other potential
hosts, while accumulation of rabies virus in the salivary glands
ensures that it is spread by bites (Krebs, J. W. et al. 1995).
Horizontally transmitted parasites which target nervous tissue
increase transmissibility by modifying the host into a suicidal
instrument of transmission.
Transmission factors determining parasitic virulence are the
spatial element in a spatial-temporal dynamic. Host density directly
determines the virulence of parasites which depend upon a single host
species (Herre 1993). Virulence may be increased when transmission
necessitates insect vectors or consumption of the primary host by
another species. Virulence varies inversely with the distance between
potential hosts; this distance is magnified when it is measured
between different species.
THE EQUILIBRIUM MODEL
It has been proposed that there is a coevolutionary arms race
between parasite and host, as the former seeks to circumvent the
defensive adaptations of the latter (Esch and Fernandez 1993). In this
view, parasitic virulence is the result of a dynamic stalemate between
host and parasite. This exemplifies the red queen hypothesis, which
predicts continued stalemate until the eventual extinction of both
species. Benton (1990) notes that the red queen hypothesis ignores the
potential for compromise in such a system.ructure and the Evolution of Virulence
in Nematiode Parasite of Fig Wasps. Science. 259:1442-1444.
Holmes, B. 1993. Evolution's neglected superstars. New Scientist.
140:30-34.
Hotez, P. J. and Pritchard, D. I. 1995. Hookworm Infection. Scientific
American. 272
Keymer, A. E. and Read, A. F. 1991. Behavioral Ecology: The Impact of
Parasitism pp. 37-61 In Parasite-Host Associations: Coexistence or
Conflict? Toft, C. A. et al. Eds. Oxford University Press. Oxford.
Krebs, C. J. 1994. Natural Regulation of Population Size pp. 322-348
In Ecology: The Experimental Analysis of Distribution and Abundance.
Harper Collins. New York.
Krebs, J. W., Wilson, M. L., and Childs, J. E. 1995. Rabies -
Epidemiology, Prevention, and Future Research. Journal of Mammalogy.
76:681-694.
Lenski, R. E. and May R M. 1994. The Evolution of Virulence in
Parasites and Pathogens: Reconciliation Between Two Competing
Hypotheses. Journal of Theoretical Biology. 169:253-265.
Milinski, M. 1990. Parasites and Host Decision-Making pp. 95-116 In
Parasitism and Host Behaviour. Barnard, C. J. and Behnke, J. M. Eds.
Taylor & Francis. London.
Read, A. F. and Harvey, P. H. 1993. The evolution of virulence theoretical framework in which the factors dft, C. A. and Aeschlimann, A. 1991. Introduction: Coexistence or
Conflict? pp. 1-12 In Parasite-Host Associations: Coexistence or
Conflict? Oxford University Press. Oxford.
Wise, L. A. (1990) Small Animal Medical Nursing pp. 226-261 In
Clinical Textbook for Veterinary Technicians, Second Edition.
McCurnin, D. M. Ed. W. B. Saunders Company. Philadelphia.